Chap. 37. Nutrient and Transport in Plants
|
37-1. |
Plants require inorganic nutrients |
|
37-2. |
How water moves through a plant |
|
37-3. |
How organic nutrients are transported |
|
37-4. |
Biological relationships |
植物需要無機養份
1
、必須有可辨識出的營養價值2
、必須無其他元素可替代3
、缺乏它則植物必亡C
,H,O→組成植株乾重之96%C
表
37.1之元素為來自土壤,N來自NH4+與NO3-,NH4+經細菌消化分解成NO3-,植物吸收NO3-,另外豆科作物的根瘤中有共生菌可吸收空氣中的N2,稱固氮作用。37-1-1. How are requirements determined?
37-1-2. What affects mineral availability?
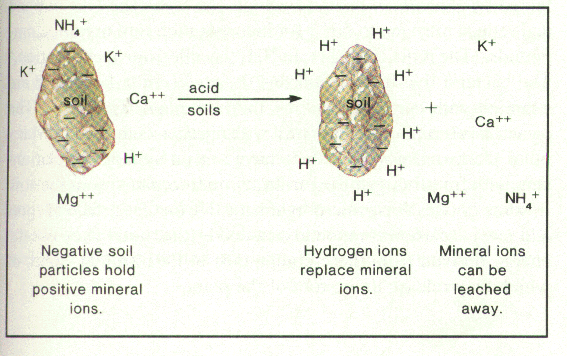
影響可吸收無機物存在量的因素?土中無機物之位置,水量及水之
pH值。pH低之水脫鹽能力高,於是根系便較難吸收到營養。土壤中(尤其是黏土及其他有機物)通常帶負電,會吸收Ca+2,K+,Mg+2等正離子,使其容易被根系所吸收,但酸性土壤中所吸收的正離子則被H+取代,此些正離子成自由態所以容易被水分溶脫而流失,於是土壤中便減少了可吸收的營養元素。然而,在高酸性的土壤中,
Al+3及Fe+3 become available,此些離子在pH>4是不可溶的,它們亦可取代Ca+2 ,K+ ,Mg+2等正離子就與H+相同。高Al+3與Fe+3是有毒性的(對植物言之)。由此可見酸性沈澱物(來自酸雨/酸雪)對植物有兩大害處:
| Certain elements are required by plants for good nutrition. Lack of an essential nutrient causes plants to die. Beneficial nutrients serve particular purposes in certain plants. |
37-1-3.
How minerals are taken in and distributed營養元素的輸送,基本上與水分相同。
植物根系吸收營養的能力是頗驚人的,通常其累積量為周圍土壤中的許多倍。在根系中某些營養元素的濃度甚至是周圍土壤中的
10000倍以上。營養元素隨著水沿木質部往上輸送,有些直達葉子,有些在中途就離開而進入那些需要營養的細胞。
營養元素如何穿越原生質厚膜? 離子態營養元素的輸送需間接利用
Adenosine Triphosphate (ATP, 三磷酸腺口票口令核甘?)的能量。植物細胞會消耗能量來取得並收藏營養元素之離子(如
N2以No3-or NH4+,P為HPO4-3,K為K+),如果根系缺乏O2或受毒害,則細胞的呼吸作用無法進行,則營養元素便無法取得。質子泵:
ATP水解產生ADP+P,此產生的能量將H+輸出至細胞外,造成電化學上的梯度,使得正電離子如K+ ,會經由蛋白質通道進入細胞,帶負電的營養離子或部份正離子則在H+ 外移之同時,經由蛋白質載體進入 (Fig.37.2)。37-1-3.1 How roots are adapted for uptake
有兩種共生方式可協助根系來取得營養元素
似共生
(semi-symbiotic relationship):植物與土中具固氮能力的微生物。 在某些草中,此些微生物住在根系的表面,接受根系所排放出來的碳化合物做能量來源進行固氮,部份產物排出為根系所吸收。極細的菌絲
許多植物均頗依賴根菌,如蘭科種子必須有根菌侵入才能發芽。有些不行光合作用的植物如水晶蘭(鹿蹄草科
, Indian pipe)即是靠其根菌來吸收臨近樹木的養分。另有些植物的根系功能頗差或完全無根,主要是由於水分與營養的供應機制有所不同,如肉食性植物如捕蠅草與毛氈苔。
附生植物
(Epiphytes)為空中植物(air plants),不長於土中,而是長在較大的植株上,靠大植株支撐,但不由大植株取得營養。有些附生植物有根可吸收空氣中的水分,有些則在葉子部份有特殊的口袋可捕捉雨水及水中的營養元素。寄生植物
(parasitic plants)如菟絲子(dodder)、內蓯蓉(broom rapes)等,其根系稱為吸根或寄生根,可穿透寄主的莖部深入至木質部與韌皮部。| Most plants are assisted in acquiring minerals by a symbiotic relationship with microorganisms and/or fungi. Some, such as predaceous or parasitic plants, have special adaptations for minerals acquisition. |
水分要由根輸送至葉,其必須經過原生質膜
(plasma membranes),以下讓我們來探討植物學家所提出的一個輸送模式。37-2-1. Water potential is critical
水勢是關鍵
水勢為水的位能,水由高位能處向低位能處流動,就像瀑布的水向下流一樣。在細胞中兩個因素決定水份穿透原生質膜的流動方向
(fig37.6),此些因素為:水份將由水壓高(具高壓力位能)處流向水壓低處。水份也會由低溶質濃度區域流向高溶質濃度區域。較高溶質濃度的區域有較低的滲透勢
(osmotic potential)水份由高滲透勢區流向低滲透勢區。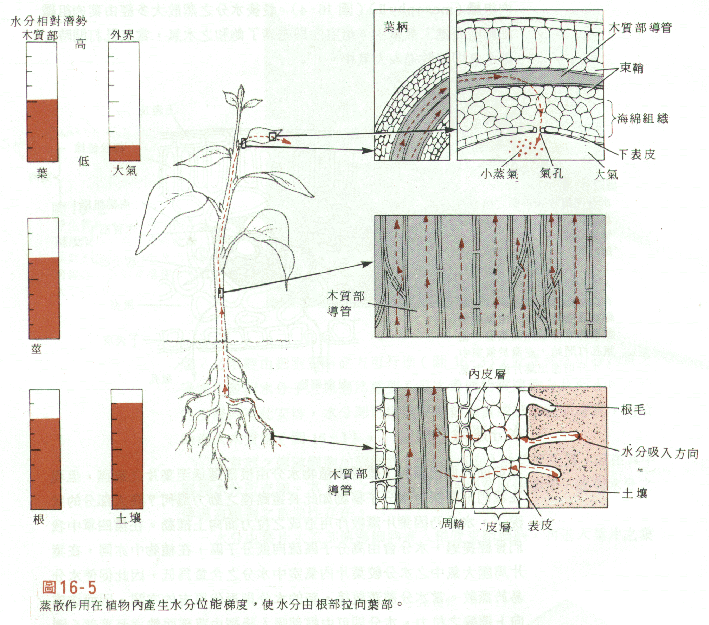
假設水分滲透入具較高溶質濃度的細胞中,細胞內水分增加所以水壓增加,當水壓(壓力位能)增加到與外界的滲透勢達到一平衡狀態時,水分即不再進入。所以說壓力位能與滲透勢為決定水勢的兩個主要因素,此兩者決定水的流向。
由於滲透所造成的壓力位能增加稱之為膨壓(turgor pressure),植物能保持飽滿即依靠此膨壓。若反向則植物成凋萎狀。
| Water moves across plasma membranes from the area of higher water potential to the area of lower potential. |
37-2-2. Xylem transports water
植物中水分與礦物質的輸送涉及由根經由木質部抵達葉子。
37-1-2.1 From the roots
由根開始
如
圖37.7所示水分由土壤中進入開花植物的根系可透過細胞中間的空隙擴散。但最終在內皮層由於有嘉氏帶,所以最後水分會進入內皮細胞(parthway A);亦可透過根毛,經由一個個細胞經過表皮及皮層再進入內皮細胞(parthway B)。不論何種路徑,水分能由土壤進入根系即代表根系中的水分比外部土壤中的水分有較低的水勢。水分進入根系的細胞產生正壓稱之為根壓
(root pressure),根壓主要在夜晚發生,常逼使木質部中的樹液向上流。根壓基本上是造成泌水現象(guttation)的原因(fig37.8)。除了根壓之外,植物要把水由根部輸送至頗高的樹上,應有其它機制配合。37-2-2.2 When is water available?
何時有水
雨後或灌溉後,重力使土壤中的水份流失,其流失速度與土壤的穿透性有關。沙土最易,黏土最難。在流失後仍存於土壤中的水分稱為田間容水量
(field capacity)。土壤中有水不表示一定可被植物吸收,不再能被植物根系吸收時的容水量稱之為永久凋萎點
(permanent wilting point)。以黏土為例,儘管仍含水分,但其被小顆粒土牢牢吸住,土中與根系中的水勢變的一樣或者前者更低,如此根系即無法由土中吸水。最理想的土壤必需含有水分且必需有空隙(根系的細胞需有氧氣進行呼吸)且必需容易讓根穿透。最好的農用土為沃土
(loams)其為10-25%黏土,其它則為1比1的砂與淤泥(silt)。| Water and minerals must enter root cells before they reach xylem. Water enters root cells because the water potential within root cells is less than that of the soil solution. |
37-1-2.3 To the leaves
直達葉子
水分進入木質部後便必需經過整個莖的高度,如美國紅杉
(red wood)的莖可能超過110公尺,此可是一個長程輸送。 圖37.9所示的內聚力-牽引力模式(cohesion-tension model)為木質部的輸送現象提出解釋。內聚力為水分子集結一起的傾向。水分子間透過氫鍵連結,由根至葉有一連續的水柱不輕易斷開,黏滯力(Adhesion)為水的另一性質可防止水柱往下滑落,主要為水柱與木質部管壁間的作用力。整個水柱向上移動則導因於牽引力
(tension),是一個負的壓力位能,因水分由葉子蒸散(transpiration)所造成。由根系吸收的水份至少有90%被蒸散出來,在一個生長季(四個月)中,一株玉米所蒸散的水分量可高達135至200公升。當植物遭受水逆境
(water stress),氣孔關閉,如此即無法吸收CO2行光合作用。所以要維持正常生長便應提供足量的水份使氣孔不致關閉。| The driving force for the ascent of water in xylem is a negative pressure potential brought about by transpiration at he leaves. |
37-2-3. Opening and closing of stomates
| When stomates open, first K+ and then water enters guard cells. Stomates open and close in response to environmental signals, and the exact mechanism is being investigated. |
有機養分如何傳輸
植物除了要能輸送水及無機的營養之外(由根至葉)還要能夠輸送有機的營養至所有需要的部位,如初生的葉子,正在製造種子及果實的花及根系等。其輸送主要在韌皮部進行。
37-3-1. Phloem transports organic nutrients
韌皮部傳輸有機營養
1679
年Malpighi觀察到將樹幹予以環剝(girdling)則在切口處會腫脹,且在樹皮的腫脹部份會有糖份累積,所以他認為樹皮(bark)應與糖份由葉轉移至根的輸送有關。環剝時木質部仍存在,所以應是樹皮中的韌皮部與此傳輸有關。使用
C14做輻射追蹤亦證實這點。另外亦發現如氨基酸,荷爾蒙及離子態微量元素亦會由葉子中移走而進入其他部位。韌皮部中的樹液,經過化學分析其主要成分為蔗糖
(sucrose),通常佔10-13%的體積。傳統上要取得樹液而不傷害植物是不可能的,但如今發現可透過蚜蟲(aphids)來進行(如Fig.37.11)。蚜蟲將其尖銳的口器(stylet)插入表皮中,再由篩管細胞抽取樹液。將蚜蟲以乙醚麻醉(anesthetized)後切去身體可取得樹液。37-3-2.
Phloem transport uses positive pressure韌皮部利用正壓進行輸送
在莖部的上下兩個地方進行取樣,利用
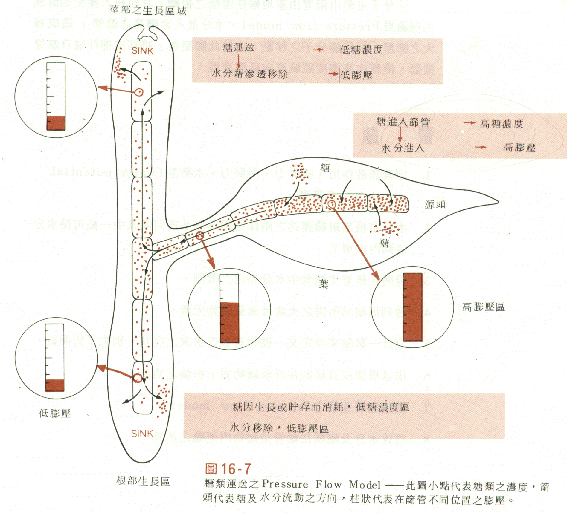
C14輻射追蹤發現在韌皮部內流動的物質其速度為60-100cm/hr亦可能高至300cm/hr。有關物質在韌皮部內的輸送目前係採壓力-流動模式來解釋 (Fig.37.12)。在生長季節,葉子行光合作用生產糖份,蔗糖被送入韌皮部,此機制與前所述之質子泵造成電化學梯度之解釋是相同的。蔗糖與
H+一起穿越細胞膜(membrane)基於濃度差。水進入篩管乃因後者有較低的滲透勢(osmotic potential)。篩管細胞進行此輸送的能量來自其伴隨細胞(companion cells)。水分的滲入在篩管內造成一個正壓,此點有別於根系之吸收。由葉子至根系,在韌皮部內有一正壓梯度
(positive pressure potential gradient),水分及蔗糖最後由根系排出。此壓力-流動模式可用下列試驗證明:
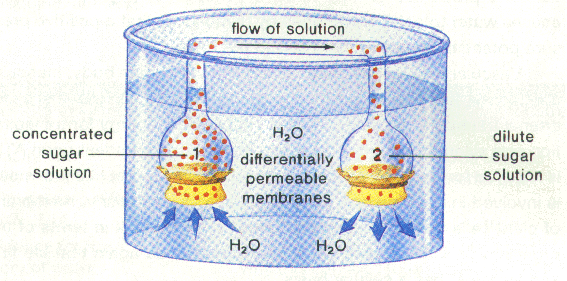
1
內為高濃度糖水,所以為低水勢H2O滲透進入1後造成其內的壓力勢提高。此「壓力勢」足夠將H2O壓至2並強制將H2O輸出,儘管2之糖濃度還高於外界溶液。在韌皮部的篩管細胞中樹液的流動,基本上是由源點
(source)至匯點(sink)的輸送,此即壓力-流動模式。在小苗中子葉
(cotyledon)部份儲存食物為蔗糖的源點,根系則為匯點。在較大植株其新生葉子可為另一匯點,此時的輸送則由鄰近行光合作用的葉片至此新生葉。結果實之時,韌皮部內的輸送變為單一方向,幾乎全往果實部輸送,極少部份至植株的其他地方,此時營養生長
(vegetative growth)變緩。韌皮部內的輸送有時可能是雙向的,目前尚不知此雙向的流動是發生在同一篩管細胞內或經由不同的篩管細胞。
| The pressure-flow model of phloem transport suggests that phloem sap can move either up or down as appropriate for the plant at a particular time in its life cycle. |
生物的關係
生物學家探究生命的化學及物理基礎,他們發現透過維管的輸送總是需要物理的力量
(physical forces),在人體心臟輸送血液產生血壓,就像機械泵將水加壓。在植物上,葉子中水分蒸散造成負壓將水由根部經木質部輸送至葉,其同時依賴水的內聚力與黏滯力。糖份主動輸送至韌皮部造成水份滲透進入篩管。此正壓勢驅使韌皮部的樹液直達根部。維管輸送需要能源。在人體,肌肉的收縮靠
ATP分解來提供能量給動脈。在植物,陽光提供能量供水分蒸散及木質部的傳輸。韌皮部的輸送仍依賴ATP做主動輸送。化學滲透(chemiosmosis)是無機物在植物原生質膜間傳輸的主動機制。生理時鐘對動、植物都有影響,人身在暗處仍能有正常的作息時間,但長時間下來此時鐘會不準而通常可透過環境的刺激而予以重調。植物的氣孔旁的保衛細胞亦同,植物沒有神經,而如何對環境刺激加以反應是目前的研究課題。重點包括如何接收,如何轉換成可影響新陳代謝的形式,反應如何發生。就人體而言,已知視網膜將光線刺激轉化為神經脈衝再由腦部接收。
儘管我們對動、植物常以器官來認知,但最終其組成還是要回溯至細胞的層次。儘管二者在外觀上有很大差別,但由細胞的觀點來看,很多機制對動、植物都是一樣的,生命的單純化
( Unity of life)說明了物種的相同起源。
